|
The efferents of both the anteroventral and posteroventral nuclei contribute to the trapezoid body, which terminates at the pontine level in the complex of the superior olives complex (SOC)(Fig 1.2). The SOC processes information about interaural delays and amplitudes. It also acts as a crossover site for spatially oriented auditory information. Receiving both ipsi- and contralateral inputs, the medial and lateral olives represent the first level of binaural convergence and play an essential role in sound localization using two localization cues: interaural time and intensity differences (ITD and IID). Some superior olivary neurons respond selectively to a certain direction of frequency modulation (Watanabe et al., 1968; Vartanjan, 1978). Furthermore, some olivary neurons were found to synchronize their activity with relatively high modulation frequencies (150 – 200 Hz) of amplitude-modulated sounds (Andreeva and Vasil’ev, 1977).ffice ffice" /> ffice" /> The outputs from medial and lateral superior olives extend to the midbrain auditory structure, the inferior colliculi (IC) (Fig 1.2), via the lateral lemniscus (LL). The LL is primarily a tract of axons ascending the brainstem. The LL however are sensitive to changes in both the timing and the amplitude of sound. The IC appears to be an integrative station as well as a switchboard. It is responsive to interaural delay and amplitude differences and may provide a spatio-topic map of the auditory environment. The IC consists of two main components: the multi-layer central nucleus, which receives most of its projections from lower auditory structures, and the dorsal part, which receives both auditory and somato-sensory input. Within the IC there are areas in which axons from different brainstem nuclei converge integrating information from the lower level (Oliver et al., 1997). The inferior colliculi obviously play an important role in sound localization since this structure contains numerous neurons sensitive to the interaural time and intensity difference (Altman, 1990; Hudspeth, 2000a; Bear et al., 2001). In response to amplitude- or frequency-modulated sounds, inferior collicular neurons are able to synchronize their firing rate with modulation frequency up to 30 – 100 Hz. This synchronization pattern becomes more robust as the stimulus carrier frequency gets closer to the characteristic frequency of a given neuron (Vartanjan, 1978). Such selectivity to particular combinations of complex stimulus parameters may result in an orthogonal representation of timing and spectral information in the IC (Langner and Schreiner, 1988). While the central nucleus is the main origin of cochleotopic projections to the thalamic level, nuclei of the dorsal part constitute multiple diffuse ascending pathways (Andersen et al., 1980; Calford and Aitkin, 1983). The most prominent pathways connecting midbrain and thalamic auditory structures, the medial geniculate bodies (MGB), are the brachia of inferior colliculi (Fig 1.2). The functions of MGB are relative intensity comparison, duration comparison and relay to cortex. The thalamus is the last relay site on the way to the cortex for almost all sensory information, including auditory, visual, and somatosensory. It is shaped like a football and has been functionally and morphologically divided into a large number of nuclei. the primary nucleus involved in the auditory pathway is the MGB, but several other nuclei are at least partially responsive to auditory stimulation. The MGB has itself been divided into three subneclei based on functional as well as morphological distinctions. These divisions are ventral (VMBG), Medial (MMGB), and dorsal (DMGB). The VMGB is thought to be primarily responsible for relaying frequency, intensity and binaural information to the cortex. The responses in the VMGB appear to be organized in a tonotopically similarly way to those in the IC. The MMGB seems to functionally be responsible for detection of the relative intensity and duration of a sound. It shows a wide range of responses to auditory stimuli. Many types of responses are present in the DMGB which appear to vary by sub-nuclei. Generally, the responses are broadly tuned, but some cells appear to respond only to complex stimuli. Other cells are multi modal, often responding to somatosensory as well as auditory stimuli (see http://serous.med.buffalo.edu/hearing/thalamus.html). The main MGB efferents project tonotopically through the acoustic radiation to the primary auditory cortex. The projections from MGB to auditory cortex preserve the segregation into tonotopic, diffuse and polysensory pathways. The tonotopic pathway goes from ICC to MGBv to the four fields of tonotopic cortex. The diffuse pathway projects from dorsal cortex of the IC to MGBd to secondary cortical areas. The polysensory pathway goes from IC paracentral nuclei to medial MGB to all cortical areas. At the mesencephalic level, there are connections between the MGB and associative thalamic nuclei: dorso-medial (DM), dorso-lateral (DL), postero-lateral (PL) and Pulvinar. These nonspecific thalamic nuclei, in turn, constitute extensive projections to frontal and parietal associative cortical areas. The DM nucleus sends efferents to Brodmann areas 8 – 12 and 45 – 47, the DL and PL nuclei to parietal areas 5 and 7, while the Pulvinar is connected to areas 39 and 40 (Andreeva et al., 1985). It should be emphasized that some auditory information circumvents the primary auditory cortex and is transferred directly to polymodal areas. However, the main MGB efferents project tonotopically through the acoustic radiation to the primary auditory cortex. [此帖子已被 zhixl 在 2005-10-31 14:29:22 编辑过] | 

 IP卡
IP卡 狗仔卡
狗仔卡 楼主
楼主
 显身卡
显身卡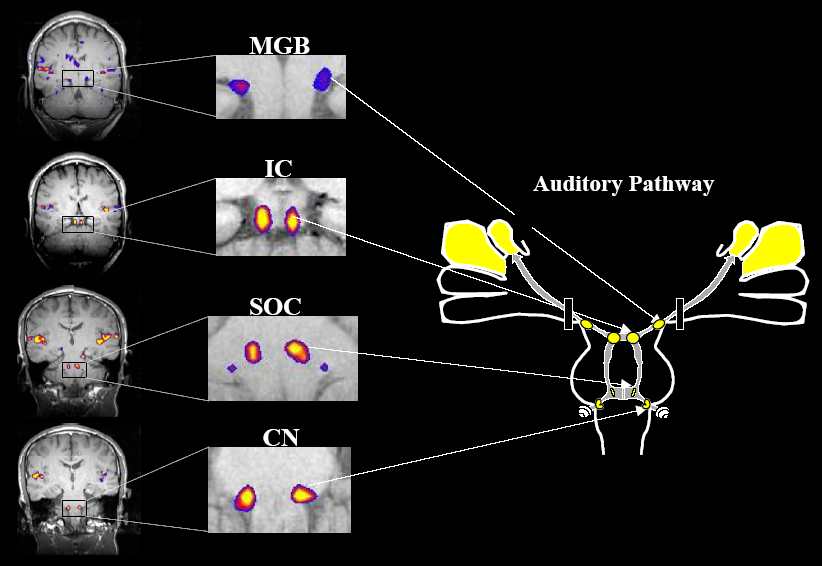

 Superb!
Superb!